
Meiosis I
Interphase
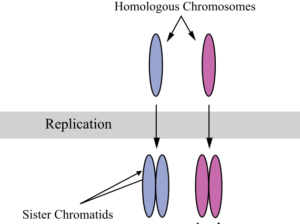
Meiosis is preceded by an interphase which is nearly identical to the interphase preceding mitosis. During interphase, the DNA of the chromosomes is replicated (during S phase). After DNA replication, each chromosome becomes composed of two identical copies (called sister chromatids) that are held together at the centromere until they are pulled apart during meiosis II (Figure 1).
Meiosis I
Meiosis is preceded by an interphase consisting of the G1, S, and G2 phases, which are nearly identical to the phases preceding mitosis. The G1 phase, which is also called the first gap phase, is the first phase of the interphase and is focused on cell growth. The S phase is the second phase of interphase, during which the DNA of the chromosomes is replicated. Finally, the G2 phase, also called the second gap phase, is the third and final phase of interphase; in this phase, the cell undergoes the final preparations for meiosis.
During DNA duplication in the S phase, each chromosome is replicated to produce two identical copies, called sister chromatids, that are held together at the centromere by cohesin proteins. Cohesin holds the chromatids together until anaphase II. The centrosomes, which are the structures that organize the microtubules of the meiotic spindle, also replicate. This prepares the cell to enter prophase I, the first meiotic phase.
Prophase I
Early in prophase I, the chromosomes can be seen clearly microscopically. As the nuclear envelope begins to break down, the proteins associated with homologous chromosomes bring the pair close to each other. The tight pairing of the homologous chromosomes is called synapsis (Figure 2). In synapsis, the genes on the chromatids of the homologous chromosomes are precisely aligned with each other. Recall that synapsis does NOT occur during mitosis.
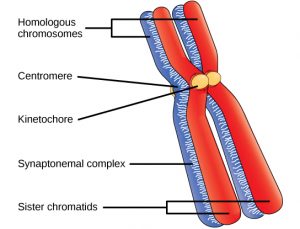
In synapsis, the genes on the chromatids of the homologous chromosomes are aligned precisely with each other. An exchange of chromosome segments between non-sister homologous chromatids occurs and is called crossing over (Figure 3). The crossover events are the first source of genetic variation produced by meiosis. A single crossover event between homologous non-sister chromatids leads to a reciprocal exchange of equivalent DNA between a maternal chromosome and a paternal chromosome. Now, when that sister chromatid is moved into a gamete, it will carry some DNA from one parent of the individual and some DNA from the other parent. The recombinant sister chromatid has a combination of maternal and paternal genes that did not exist before the crossover.
Prometaphase I
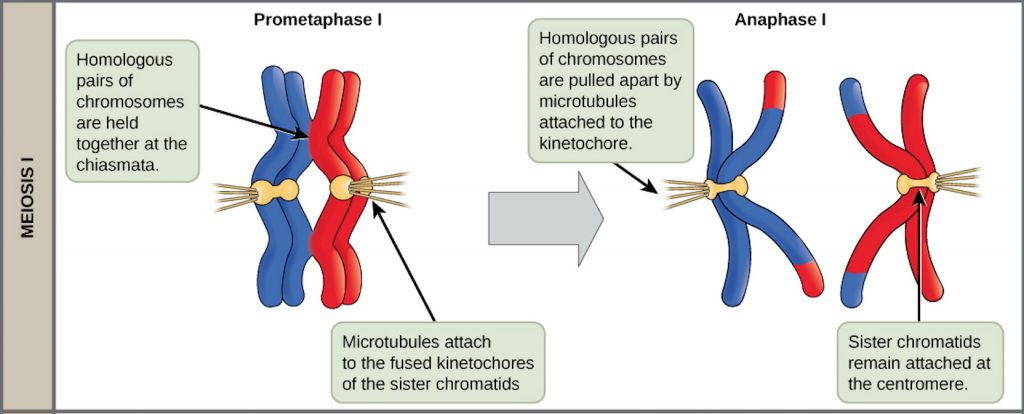
The key event in prometaphase I is the attachment of the spindle fiber microtubules to the kinetochore proteins at the centromeres. Kinetochore proteins are multiprotein complexes that bind the centromeres of a chromosome to the microtubules of the mitotic spindle. Microtubules grow from centrosomes placed at opposite poles of the cell. The microtubules move toward the middle of the cell and attach to one of the two fused homologous chromosomes. The microtubules attach at each chromosomes’ kinetochores. With each member of the homologous pair attached to opposite poles of the cell, in the next phase, the microtubules can pull the homologous pair apart. A spindle fiber that has attached to a kinetochore is called a kinetochore microtubule. At the end of prometaphase I, each tetrad is attached to microtubules from both poles, with one homologous chromosome facing each pole. The homologous chromosomes are still held together at chiasmata. In addition, the nuclear membrane has broken down entirely.
Metaphase I
During metaphase I, the homologous chromosomes are arranged in the center of the cell with the kinetochores facing opposite poles. The orientation of each pair of homologous chromosomes at the center of the cell is random. This randomness, called independent assortment, is the physical basis for the generation of the second form of genetic variation in offspring (Figure 5). Consider that the homologous chromosomes of a sexually reproducing organism are originally inherited as two separate sets, one from each parent in the egg and the sperm. Using humans as an example, one set of 23 chromosomes is present in the egg donated by the mother. The father provides the other set of 23 chromosomes in the sperm that fertilizes the egg. In metaphase I, these pairs line up at the midway point between the two poles of the cell. Because there is an equal chance that a microtubule fiber will encounter a maternally or paternally inherited chromosome, the arrangement of the tetrads at the metaphase plate is random. Any maternally inherited chromosome may face either pole. Any paternally inherited chromosome may also face either pole. The orientation of each tetrad is independent of the orientation of the other 22 tetrads.
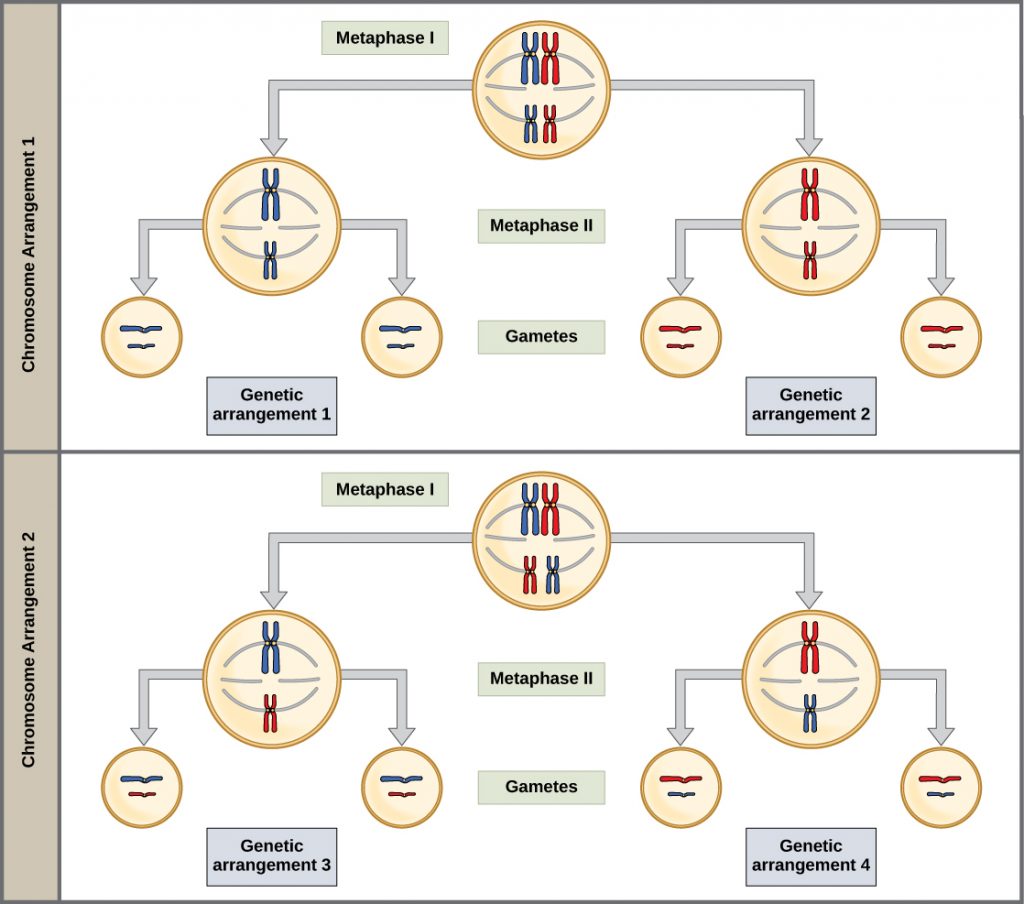
To summarize the genetic consequences of meiosis I: the maternal and paternal genes are recombined by crossover events occurring on each homologous pair during prophase I; in addition, the random assortment of tetrads at metaphase produces a unique combination of maternal and paternal chromosomes that will make their way into the gametes.
Anaphase I
In anaphase I, the microtubules pull the linked chromosomes apart. The sister chromatids remain tightly bound together at the centromere. The chiasmata are broken in anaphase I as the microtubules attached to the fused kinetochores pull the homologous chromosomes apart (Figure 4).
Telophase I and Cytokinesis I
In telophase, the separated chromosomes arrive at opposite poles. The remainder of the typical telophase events may or may not occur, depending on the species. In some organisms, the chromosomes decondense and nuclear envelopes form around the chromatids in telophase I. In other organisms, cytokinesis—the physical separation of the cytoplasmic components into two daughter cells—occurs without reformation of the nuclei. In nearly all species of animals and some fungi, cytokinesis separates the cell contents via a cleavage furrow (constriction of the actin ring that leads to cytoplasmic division). In plants, a cell plate is formed during cell cytokinesis by Golgi vesicles fusing at the metaphase plate. This cell plate will ultimately lead to the formation of cell walls that separate the two daughter cells.
Two haploid cells are the end result of the first meiotic division. The cells are haploid because at each pole, there is just one of each pair of the homologous chromosomes. Therefore, only one full set of the chromosomes is present. This is why the cells are considered haploid—there is only one chromosome set, even though each homolog still consists of two sister chromatids. Recall that sister chromatids are merely duplicates of one of the two homologous chromosomes (except for changes that occurred during crossing over). In meiosis II, these two sister chromatids will separate, creating four haploid daughter cells.
Summary of Meiosis I
The chromosomes are copied during interphase (prior to meiosis I). This forms two identical sister chromatids that are attached together at the centromere. During prophase I, crossing over introduces genetic variation by swapping pieces of homologous chromosomes. Additional genetic variation is introduced by independent assortment, which takes into account how the homologous chromosomes line up during metaphase I. At the end of meiosis I, two haploid cells (where each chromosome still consists of two sister chromatids) are produced.
References
Unless otherwise noted, images on this page are licensed under CC-BY 4.0 by OpenStax.
OpenStax, Biology. OpenStax CNX. January 2, 2017 http://cnx.org/contents/s8Hh0oOc@9.10:1Q8z96mT@4/Meiosis

{kind=link}