
Response to the signal
Ligands which can enter the cell and bind to internal receptors are able to directly affect the cell’s DNA and protein-producing machinery. Ligands which can not enter the cell bind to receptors in the plasma membrane and use signal transduction pathways to produce a variety of effects. The results of signaling pathways are extremely varied and depend on the type of cell involved as well as the external and internal conditions. A small sampling of responses is described below.
Gene Expression
Some signal transduction pathways regulate the production of mRNA. Others regulate the synthesis of proteins from mRNA by ribosomes. Typically these pathways increase gene expression so that more of a specific protein is produced from a gene, but some pathways do decrease gene expression.
An example of a protein that regulates translation in the nucleus is the MAP kinase ERK. ERK is activated in a phosphorylation cascade when epidermal growth factor (EGF) binds the EGF receptor. Upon phosphorylation, ERK enters the nucleus and activates a protein kinase that, in turn, regulates protein translation (Figure 1).
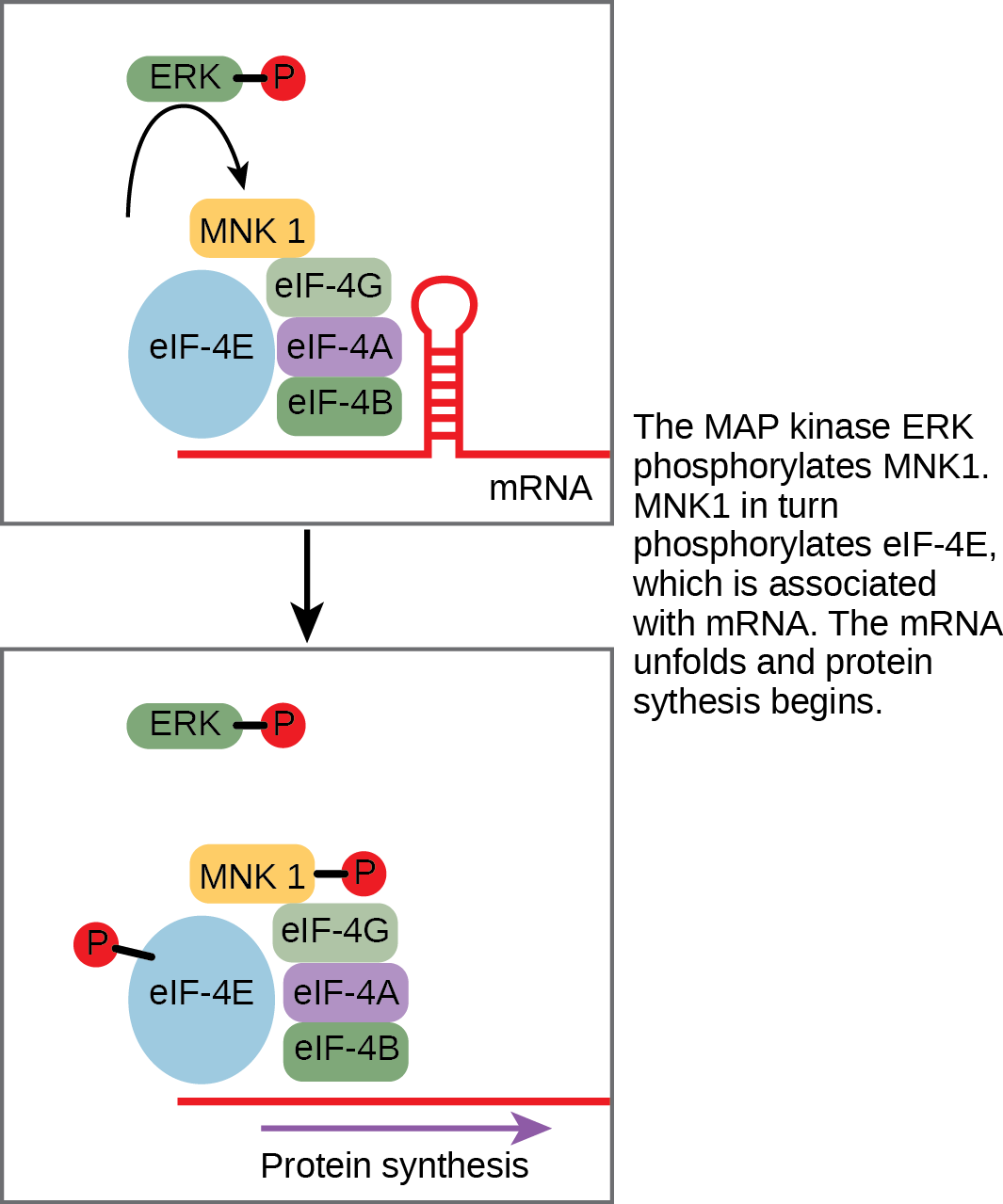
Increase in Cellular Metabolism
The result of another signaling pathway affects muscle cells. The activation of β-adrenergic receptors in muscle cells by adrenaline leads to an increase in cyclic AMP (cAMP) inside the cell. Also known as epinephrine, adrenaline is a hormone (produced by the adrenal gland attached to the kidney) that readies the body for short-term emergencies. Cyclic AMP activates PKA (protein kinase A), which in turn phosphorylates two enzymes. The first enzyme promotes the degradation of glycogen by activating intermediate glycogen phosphorylase kinase (GPK) that in turn activates glycogen phosphorylase (GP) that catabolizes glycogen into glucose. (Recall that your body converts excess glucose to glycogen for short-term storage. When energy is needed, glycogen is quickly reconverted to glucose.) Phosphorylation of the second enzyme, glycogen synthase (GS), inhibits its ability to form glycogen from glucose. In this manner, a muscle cell obtains a ready pool of glucose by activating its formation via glycogen degradation and by inhibiting the use of glucose to form glycogen, thus preventing a futile cycle of glycogen degradation and synthesis. The glucose is then available for use by the muscle cell in response to a sudden surge of adrenaline—the “fight or flight” reflex.
Cell Growth
Cell signaling pathways also play a major role in cell division. Cells do not normally divide unless they are stimulated by signals from other cells. The ligands that promote cell growth are called growth factors. Most growth factors bind to cell-surface receptors that are linked to tyrosine kinases. These cell-surface receptors are called receptor tyrosine kinases (RTKs). Activation of RTKs initiates a signaling pathway that includes a G-protein called RAS, which activates the MAP kinase pathway described earlier. The enzyme MAP kinase then stimulates the expression of proteins that interact with other cellular components to initiate cell division.
Cell Death
When a cell is damaged, superfluous, or potentially dangerous to an organism, a cell can initiate a mechanism to trigger programmed cell death, or apoptosis. Apoptosis allows a cell to die in a controlled manner that prevents the release of potentially damaging molecules from inside the cell. There are many internal checkpoints that monitor a cell’s health; if abnormalities are observed, a cell can spontaneously initiate the process of apoptosis. However, in some cases, such as a viral infection or uncontrolled cell division due to cancer, the cell’s normal checks and balances fail. External signaling can also initiate apoptosis. For example, most normal animal cells have receptors that interact with the extracellular matrix, a network of glycoproteins that provides structural support for cells in an organism. The binding of cellular receptors to the extracellular matrix initiates a signaling cascade within the cell. However, if the cell moves away from the extracellular matrix, the signaling ceases, and the cell undergoes apoptosis. This system keeps cells from traveling through the body and proliferating out of control, as happens with tumor cells that metastasize.
Another example of external signaling that leads to apoptosis occurs in T-cell development. T-cells are immune cells that bind to foreign macromolecules and particles, and target them for destruction by the immune system. Normally, T-cells do not target “self” proteins (those of their own organism), a process that can lead to autoimmune diseases. In order to develop the ability to discriminate between self and non-self, immature T-cells undergo screening to determine whether they bind to so-called self proteins. If the T-cell receptor binds to self proteins, the cell initiates apoptosis to remove the potentially dangerous cell.
Apoptosis is also essential for normal embryological development. In vertebrates, for example, early stages of development include the formation of web-like tissue between individual fingers and toes (Figure 2). During the course of normal development, these unneeded cells must be eliminated, enabling fully separated fingers and toes to form. A cell signaling mechanism triggers apoptosis, which destroys the cells between the developing digits.
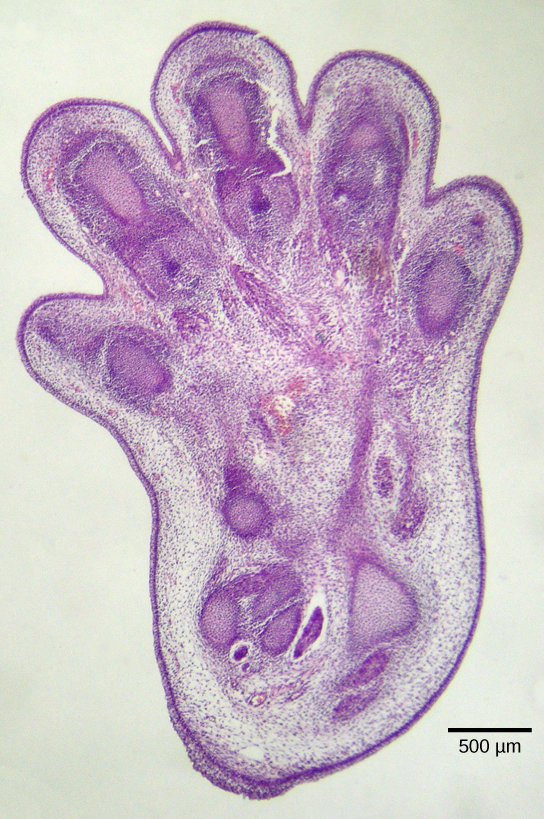
Termination of the Signal Cascade
The aberrant signaling often seen in tumor cells is proof that the termination of a signal at the appropriate time can be just as important as the initiation of a signal. One method of stopping a specific signal is to degrade the ligand or remove it so that it can no longer access its receptor. One reason that hydrophobic hormones like estrogen and testosterone trigger long-lasting events is because they bind carrier proteins. These proteins allow the insoluble molecules to be soluble in blood, but they also protect the hormones from degradation by circulating enzymes.
Inside the cell, many different enzymes reverse the cellular modifications that result from signaling cascades. For example, phosphatases are enzymes that remove the phosphate group attached to proteins by kinases in a process called dephosphorylation. Cyclic AMP (cAMP) is degraded into AMP by phosphodiesterase, and the release of calcium stores is reversed by the Ca2+ pumps that are located in the external and internal membranes of the cell.
References
OpenStax, Biology. OpenStax CNX. October 13, 2017. https://cnx.org/contents/GFy_h8cu@10.118:yQ4MP9JZ@7/Response-to-the-Signal
